
Source: http://www.tulane.edu/~wiser/protozoology/notes/INTRO.html
Mitä Alkueläimet?
Selvittämisessä Kreikan juuret tulokset määrittelyssä alkueläimiä kuin “ensimmäinen” (proto) ‘eläimillä’ (Zoa). Vaikka molekyylitason fylogeneettisissä tutkimukset osoittavat, että alkueläimet ovat yksi varhaisimmista haarautumisen eukaryooteissa (ks fylogeneettiseen puu), tällainen määritelmä ei tarjoa paljon kuvailevaa tietoa. Alkueläimet ei ole helppo määritellä, koska ne ovat erilaisia ja ne ovat usein vain kaukaista sukua toisilleen. Johtuen äärimmäisen monimuotoisuutta alkueläimet ainoa yhteinen piirre kaikille alkueläimet on, että ne ovat yksisoluisia eukaryoottisia mikro-organismeja. Alkueläimet hallussaan tyypillinen eukaryoottisia organellit ja yleensä ilmentävät tyypillisiä muita eukaryoottisolut. Esimerkiksi kalvoon sitoutunut ytimen, joka sisältää kromosomien esiintyy kaikissa alkueläin- lajeja. Kuitenkin monissa alkueläin lajien joitakin soluelimiin voi olla poissa, tai jotka ovat morfologisesti tai toiminnallisesti erilainen kuin muualla eukaryooteissa. Lisäksi monet alkueläimet ovat soluelimiin jotka ovat ainutlaatuisia tietylle alkueläimet.
Kysymykset voidaan kysyä suhteen määrittelyssä alkueläimet ovat: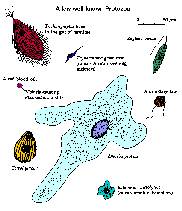
- Miltä he näyttävät?
- Kuinka suuri ne ovat?
- Missä he asuvat?
- Miten ne liikkuvat?
- Miten ne lisääntymään?
| Alkueläin koot | |
| Organismi |
Koko |
| Plasmodium merotsoiittien | 2-5 um |
| Paramecium | 200-500 nm |
| Spirostomum ambiguum | 3 mm |
| Nummulites (kivettyneet) | 7-13 cm |
| Symbioottinen yhdistykset | |
| Kommensalismi | Tarkoittaa vuorovaikutus, joka on hyödyllistä yksi organismi, mutta ei ole vaikutusta muihin organismi. Esimerkiksi monet alkueläimet elää ruoansulatuskanavan toisen organismin vahingoittamatta sitä. Nämä commensals ovat usein scavengers tai saalistajat, jotka hyödyntävät runsaasti ravinteita tai bakteerikanta, jonka isäntäorganismi. |
| Mutualismi | Tarkoittaa erityinen muoto kommensalismi jossa molemmat organismit saavat jotain hyötyä ja ovat riippuvaisia toisistaan. Klassinen esimerkki mutualismin on alkueläin Trichonympha löytyy suolistossa termiittejä. Trichonympha, avustuksella symbioosissa bakteereja, sulattaa puu- hiukkasten (so selluloosa) nautittavaksi termiitti. |
| Loisiminen | Tarkoittaa suhde, jossa yksi organismi (loinen) hyödyt kustannuksella muiden organismin (isäntä). Yleensä tämä isäntä kustannuksella merkitsee sitä, että loinen vie makromolekyylien isännältä ja vapauttaa muut vastaanottavaan. Joissakin tapauksissa loissairaus on liian haitallisia isäntä ja viitataan olevan patogeenisiä. Nämä patogeeniset alkueläimet ovat ensisijainen painopiste kurssin. |
Liikkuvuus ja solun tukirangan
| Vaadittavat liikkuvuuden | |
| Mekanismi | Subgroup |
| amoeboid liike | amebas |
| siimoja | siimalevistä |
| värekarvat | ripsieläimiä |
| luisto liikkuvuus | Sporozoa |
Varhaisimmat havainnot alkueläimet huomattava niiden liikkuvuutta. Tämä liikkuvuus on johtanut niiden luokitteleminen eläinten, jotka erotettavissa liikkumattomia “kasveja”. Kuitenkin, liikkuvuuteen ei ole universaali piirre alkueläimet ja eri alkueläimet käyttää erilaisia mekanismeja niiden liikkeen (taulukko). Itse asiassa, alkueläimiä alun perin luokitusta osittain niiden mekanismia liikkuvuuteen (ks taksonomia).
Värekarvat ja siimoja ovat subsellulaarisia rakenteita, jotka kuljettamaan alkueläimet läpi virtaavan väliaineen. Siimoja ovat pitkiä piiska kaltaisia rakenteita, jotka kuljettamaan organismin seurauksena aaltomaisen syke etenee läpi niiden pituus. Flagelloitu- alkueläimet tyypillisesti yksi tai muutama siimoja per organismi. Sen sijaan Värekarvallinen alkueläimet ovat yleensä peitetty riviä lukuisia värekarvojen. Lyöntiä Näiden värekarvojen ovat yhteensopivia ja toimivat kuten airot kuljettamaan organismin. Värekarvojen ja siimoja voi myös auttaa hankinnassa ruokaa, lisääntymiseen ja muita toimintoja. Värekarvojen ja siimoja koostuvat saman proteiinin osia ja ovat itse asiassa vastaavat rakenteet. Molemmat ovat kalvoon sitoutunut rihmamaisia ennusteet solusta. Hehkulangan, tunnettu axoneme, koostuu sarja yhdensuuntaisia mikrotubulusten, tyypillisesti osoittaen “9 + 2 ‘järjestelyä. Movement syntyy, kun mikrotubulusten liukuvat toistensa ohi. Voima, joka välittää tämä liukuva liike syntyy nimisen proteiinin dynein. Dyneins ovat “moottori proteiineja,” jotka muuttavat kemiallista energiaa vapautuu ATP hydrolyysin osaksi mekaanista energiaa. Mikrotubulukset ovat solun tukirangan tekijöitä, jotka myös tärkeitä rooleja solun muotoon ja ovat tärkeä osa sukkularihmaston.
Toisin kuin uima näytteillä siimaleviä ja ripsieläimiä, ameba ovat alkueläimet indeksoivien pitkin vankka kasvualusta on muoti kutsutaan ameboid liike “. Ameba työntyy ulos pseudopodium tai väärä jalka, solusta elin. Pseudopodium sitten kiinnittyy alustaan ja sitten vetää muun solun elin eteenpäin. Voima mukana tässä liikkeessä syntyy toinen cytoskeletal järjestelmä, joka koostuu aktiini ja myosiinin. Aktiini muodoissa pitkä filamentit, joka tunnetaan myös nimellä microfilaments, ja myosiinin on moottori proteiini, joka liikkuu pitkin microfilaments käytettäessä ATP riippuvaisella tavalla. Lihasten supistumisen on toinen esimerkki joukkojen muodostamista kautta aktiini-myosiinin sytoskeletaalisiin elementtejä. Eräässä mekanistinen mielessä, fagosytoosin on eräänlaista ameboid liikkeen mukana myös microfilaments. Tässä tapauksessa valejalka on laajennettu surround hiukkasen nautitaan. Fuusio valejalka kanssa solun elin tuloksia sisäistämisen hiukkasen sisällä vakuolin.
Apicomplexa myös ryömiä pitkin kasvualusta, mutta eri mekanismilla kuin ameba. Mekanismia tämän niin kutsutun “liukuvat motiliteettia” alkavat vasta ymmärtää ja luultavasti liittyy sekä hehkulangan ja mikrotubulusten perustuvat cytoskeletal järjestelmiä. Apicomplexa myös näytteille solunsisäisiä muotoja ja hyökkäys isäntäsolun liittyy myös tämän luisto liikkuvuutta. (Katso myös keskustelu isäntäsolun hyökkäystä malaria loinen.)
Cellular liikkuvuuteen liittyy voima sukupolvi joko mikrotubuleihin perustuva solun tukirangan elementit tai microfilamentti-pohjainen sytoskeletaalisiin elementtejä. Tämä pätee alkueläimet ja muiden eukaryoottien. Osallistuminen mikrotubulusten ja mikrofila- sekä solujen muoto ja solujen liikkumisen nämä subsellulaarisista rakenteita enemmän analoginen tuki- ja liikuntaelimistön.
Jäljentäminen
| Vaadittavat Lisääntyminen | |
| Suvuton | Binary fissio Useita fissioita Orastava |
| Seksuaalinen | Fuusio sukusolujen Konjugaatio |
Alkueläimet, kuten kaikki muutkin organismit, jäljentää. Yleisin lisääntymisen alkueläimessä on suvuton binary fissio. Toisin sanoen, yksi organismi jakaa kahteen yhtä suureen organismeihin. Pienin muutoksin tämän binary fissio, nimeltään orastava, on kun yksi vastaperustetun solut on pienempi kuin muut. Tyypillisesti suurempi solu on nimeltään äiti ja pienempi on tytär. Jotkut alkueläimet muodostavat intrasellulaarisen alkuunsa ja olennaisesti synnyttää. Toinen variaatio binary fissio on moninkertainen fissio tai segmentointia. Tässä tilanteessa on käyty useita ydin- replikointi tapahtuu ilman sytokineesiin. Tämä Monitumaiset solu sitten muodostaa useita jälkeläisiä samanaikaisesti.
Monet alkueläimet näytteille sukupuolisen lisääntymisen lisäksi suvutonta muotoihin lisääntymiseen. Tämä seksuaalinen lisääntyminen voi liittyä tuotantoa ja fuusio sukusolujen prosesseissa samanlainen eliöt. Ciliophora tehdään konjugaatio, jossa vastakkainen parittelun tyypit pariksi vaihtamaan ja geneettistä materiaalia (eli DNA). Joskus seksuaalinen lisääntyminen on pakollinen vaihe elinkaaren, kun taas muissa tapauksissa organismi voi lisääntyä suvuttomasti kanssa satunnaisesti kierros sukupuolisen lisääntymisen.
Yhteenvetona, alkueläimet ovat yksisoluisia eukaryoottisia mikro-organismeja. Kuitenkin määrä monimuotoisuuden kannalta morfologia, koko ja elämäntavat näytteille alkueläimet tekee vaikeaksi kehittää tarkempaa määritelmää. Heidän pitkä evoluution historiassa (ks fylogeneettiseen puu) johtuu suuri osa tätä monimuotoisuutta. Kuitenkin alkueläimet tehdä näytteille ominaisuuksia yhteisiä kaikille eukaryooteissa. (Linkki sarjan powerpoint esityksiä eukaryoottinen solubiologian.)
| Alkuperäinen eukaryoottisolujen |
| Evoluution hypätä prokaryooteissa ja eukaryooteissa on merkittävä ja on arviolta 5000 geenejä löytyy eukaryooteissa, joita ei löydy prokaryooteissa (1). Ensimmäinen eukaryoottiorganismeja todennäköisesti muistuttivat mitä me nyt luokitella alkueläimet. Kuitenkin luultavasti ole suoria jälkeläisiä varhaisen eukaryoottien tällä hetkellä olemassa ja polku prokaryooteissa ja eukaryooteissa on todennäköisesti täynnä useita puuttuvia yhteyksiä. Kaksi tärkeimmistä tapahtumista kehitystä eukaryoottien ovat luultavasti kehittäminen sisäisen solun tukirangan järjestelmän ja phagotrophy. Merkittävä ero eukaryoottien ja prokaryooteissa on jäykkä mureiinin tai peptidoglykaania, soluseinä bakteerien. Vaikka monet eukaryooteissa osoittavat solun seinät, ne ovat pohjimmiltaan erilaisia kuin bakteerien ja todennäköisesti peräisin toissijaisesti muodostamisen jälkeen ja eukaryooteissa. Esimerkiksi soluseinien kasvisolut koostuu pääasiassa selluloosasta ja sienten soluseinien ja kysta seinät monissa alkueläimet koostuvat pääasiassa kitiinin. Jäykkyyttä ja lujuutta tarjoamat soluseinien suojaa soluja vastaan mekaanista rasitusta ja antaa osmoottisen vakautta. Siten samanaikainen menetys Mureiini soluseinän alussa eukaryootti oli kehittää endocytoskeletal perustuvat aktiini ja tubuliinin kompensoida suhteellisen haurauden alasti solukalvon. Lisäksi alussa eukaryooteissa todennäköisesti kehittynyt on benttisen ympäristössä ja erityisesti tiheässä meren mikrobien matot. Orgaaninen liuenneet aineet erittyvät bakteerien ja korkea suolapitoisuus tällainen ympäristö olisi ollut osmoprotective vaikutus. Tiheä mikrobien matot tarjoaisi myös ympäristön, jossa saalis oli erittäin runsas ja fagosytoosin olisi voinut kehittyä ilman tehokkaita solujen liikkuvuutta. Esi eukaryoottiin luultavasti kiinni saalis kautta solun pinnalla N-glykoproteiinien joka kehittyi menettämisen jälkeen Mureiini soluseinän ja sitten hajotetaan niitä ulkoisesti erittämällä hydrolyyttisiä entsyymejä. Tehdä prosessista tehokkaamman primitiivinen saalistajat voisi olla asteittain ympäröi saaliinsa kunnes saalis oli täysin ajautuivat isäntä kalvoja ruuansulatusta toteutetaan sisäisesti fagosomeihin. Tämä endosyyttisissä mekanismi edellyttäisi myös, että luodaan exocytic mekanismi säilyttää pinta solukalvon. Siten kehitystä fagosytoosin paitsi sisälly laajennus-toiminto aktiinisytoskeletonin mutta myös ajoi kehittämistä kalvon kaupan ja subsellulaaristen myös ydin. Kehitys phagotrophy ja endomembranes paitsi avannut uuden tavan syöminen, mutta myös sallittua endosymbiosis ja orjuuttaminen mitokondrioita. Varhainen eukaryootti luultavasti nielaisi alfa-proteobacteria ja muodostivat symbioottinen suhde, joka kehittyi yhtäläistämiseltä aerobisen aineenvaihdunnan tästä organismista ja kehitystä mitokondrioissa. Tämä orjuuttaminen mitokondrioissa oli todennäköisesti toinen keskeinen tapahtuma kehittyminen eukaryoottien koska säilynyt eukaryooteissa johdetaan mitokondrion sisältävästä esi-isät. Säilynyt anaerobiset eukaryooteissa ilman mitokondrioita joko organelles johdettu mitokondrioita, kuten mitosomes ja hydrogensomes, tai ainakin ydin- geenit koodaavat jäännöksen mitokondrioiden geenien. Oletettavasti amitochondrial eukaryoottisia esi oli epäedulliseen kilpailuasemaan eikä selviytyä pitkällä aikavälillä. Myöhemmin cyanbacterium oli orjuuttama kantaisä kasvien ja kehittynyt kloroplastissa ja samalla nämä kasvien edeltäjät olivat nielaisi ja orjuuttanut muodostaa plastideihin monissa muut levät ja alkueläimet. Solun tukirangan jatkoi kehittyä ja laajentaa sen tehtävät ja erityisesti alkoi rooli solun liikkuvuuteen. Selkeimmin tubuliinin tuli keskeinen osa siimoja ja myöhemmin myosiinin kehittynyt jotta kehittämiseen ameboid liikkeen ja luiston liikkuvuutta. Tämä liikkuvuus sallittu varhaisimmat eukaryooteissa laajentua muihin markkinaraon (esim. Planktonisia, maanpäällinen) ja näin ollen tämä aerobinen, phagotrophic ja flagelloitu- esi eukaryootti voisi säteillä osaksi monipuolisista alkueläimet, ja lopulta toinen eukaryooteissa, että tällä hetkellä on olemassa. Cavalier-Smith T (2009) Saalistus ja eukaryootti solu alkuperää a coevolutionary näkökulmasta. Int. J. Biochem. Cell Biol. 41: 307-322. |
Taksonomia
Taksonomian tai systematiikka, on tiede nimeäminen ja luokittelussa organismeja. Lisäksi määrittämällä hierarkkinen taksonomisen luokittelun, systematiikka myös yrittää sijoittaa organismien ryhmiin kuvastavat kehittyvä suhteita tai phylogenies. Kuitenkin taksonominen kriteerit ovat usein mielivaltaisia ja taksonomia on aina muuttumassa heijastamaan uusia löytöjä ja tulkintoja. Lisäksi utilitaristinen ominaisuuksia, kuten sairauden tyypin, isäntäkirjo ja maantieteellinen jakauma, käytetään usein systematiikkaa patogeenisten mikro-organismien. Tämä pätee erityisesti alkueläin taksonomian. Lisäksi on jonkin verran keskustelua yleisfilosofia luokituksessa alkueläimet ja niiden väliset suhteet monet alkueläin ryhmien ei tunneta (ks D. J. Patterson Am. Nat. 154, S96-124). Luokittelu alkueläimet hankaloittaa edelleen alkuperäisen määritelmän alkueläimet ovat yksisoluisia heterotrofeiksi. Nyt on tunnustettu, että alkueläimen (tai protisteja) käyttää useita ravitsemuksellisia strategioita eikä sitä voida pitää yksinkertaisesti joko kasvi-like (autotroph) tai eläimen kaltainen (heterotrofiksi). Täten termi alkueläimet ei voida pitää todellisena taksonomiseen ryhmään. Huolimatta siitä, että sana alkueläimet sinänsä ei ole enää oikea taksonominen nimi, se on silti käyttökelpoinen ja toimiva aikavälillä.
Historiallisesti alkueläimet jaettiin neljään suureen ryhmään: ameba, siimalevät, ripsieläimet ja Sporozoa. Tuntomerkkejä ryhmien välillä perustui liikkuvuuteen (ts ameboid, siimoja, cilia). Sporozoa olivat heterogeeninen ryhmä, joka tuotti itiöitä aikana yhdessä vaiheessa elinkaarensa ja osoitti “luisto” liikkuvuutta. Tällainen luokitusjärjestelmä on varsin mielivaltainen ja ei välttämättä heijasta todellista evoluution suhteita organismeja. Eräs ongelma käyttäen liikkuvuutta kuin taksonomisen kriteereistä on, että monet alkueläimet käyttävät erityyppisiä liikkuvuuden eri vaiheissa niiden elinkaaren ajan. Esimerkiksi Naegleria esiintyy käytettäessä ameba muodossa, kun ruoka on runsasta ja muuttuu ruoskia kun ruoka on poissa. Yleensä ameba ovat heterogeeninen ryhmä ja kaikki todennäköisesti dervived alkaen siimalevät. Näistä neljä alkuperäistä alkueläin ryhmää vain ripsieläimiä pidetään edelleen voimassa luokitusjärjestelmän mukaan.
Alku 1960 elektronimikroskoopilla käytettiin tunnistamaan Mikroskooppitutkimus joita voitaisiin käyttää kriteereinä ryhmittelyä alkueläimet. Monissa tapauksissa morfologia johtaa luokituksen mikä asettaa organisims osaksi monophyletic ryhmiin. Monophyletic tarkoittaa sitä, että kaikki organismit, jotka ryhmä todennäköisesti peräisin yhteisestä esi-isästä. Esimerkiksi monet alkueläimet aiemmin nimeltään Sporozoa hallussaan subcellular rakenteita, yhdessä kutsutaan apikaalisella soluelimiin, ja muodostavat nyt monophyletic ryhmä nimeltä Apicomplexa. Kuitenkin subsellulaarisista rakenteet ja metaboliareitteihin voi kadota joillakin suvusta ja sijoittamalla ne jälkeläiset voivat olla problamatic.
1980-luvulla ja jatkuu siihen asti hetkellä molekyylitekniikkaa joita sovelletaan taksonomian. Mahdolliset evoluution historia ja suhteita voidaan saada vertaamalla DNA: n tai proteiinisekvenssien. Molecular sekvenssi data on vahvistanut phylogenies perustuu muihin kriteereihin, ratkaistaan joitakin keskusteluja, ja johti muutamia yllätyksiä. Esimerkiksi molekyyli tiedot vahvistavat, että Apicomplexa ovat monophyletic, ja lisäksi, osoittaa, että ne liittyvät ripsieläimiä ja panssarisiimalevät. Nämä kolme ryhmää ovat nyt yhdistetty suurempaan monophyletic ryhmä nimeltä alveolata. Tämä suhde oli aiemmin epäilty ja nimi on viittaus morfologisten rakenteiden tunnetaan alveolaarinen pusseja. Mutta käyttö yksittäisten geenien voi olla epäluotettava keino määrittää evoluution suhteita, etenkin etäistä sukua organismeja, ja molekyylitason tietoja olisi tulkittava varoen.
Koska 1980-luvun puolivälissä luokittelu protists on ollut muutostilassa ja kärsimään filosofisen ristiriitoja. Yhtäältä on olemassa joitakin argumentti pitoelementtien Bütschlian perustuva järjestelmä liikkuvuuteen koska sen tuntemus ja yksinkertaisuus. Nämä järjestelmät ovat usein ristiriidassa fylogeneettiseen tietoja. Lisäksi hierarkioita ja joukkoon perinteisen Linnaean systematiikka (eli pääjakso, luokka, järjestystä, perhe, suku, laji) ei aina sovi hyvin mikro-organismeja, ja se on usein vaikea päättää, mikä hierarchial taso on sopivin mitään erityistä alkueläin ryhmään.
Aiemmin monet taksonominen järjestelmät ovat määritelleet viisi valtakunnat elämän: prokaryooteissa (bakteerit), protists, kasveja, sieniä, ja eläimiä. Näissä järjestelmissä alkueläimet ovat osa Protista yhdessä yksisoluisia leviä, piileviä, oomycetes ja limasieniin. Kuitenkin on aina ollut tyytymättömyyttä alkueläin ryhmään. Tämä johtuu osittain protista määritellään osittain negatiivisena kriteerinä. Toisin sanoen, organismit, jotka eivät sovi muut neljä valtakunnat ovat automaattisesti mukana protista. Lisäksi jotkut protists ovat fylogeneettisesti läheistä sukua toiselle kolme eukaryoottiset valtakunnat kuin muille protists, ja näin protists ovat selvästi polyphyletic.
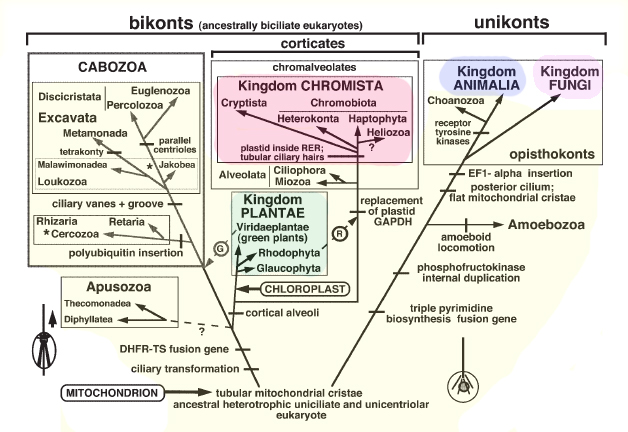
Cavalier-Smith ehdottaa viittä eukaryoottinen valtakunnat koostuu pohjapinta, ja siten paraphyletic, valtakunnan alkueläimet ja neljä johdettu valtakunnat: Animalia, sienet, Plantae, ja Chromista (kuva). Jotkut entiset protista sisältyvät nyt johdettu kuningaskunnat että ne ovat läheistä sukua jolloin valtakunnan alkueläimet tulossa monophyletic. Kuitenkin alveolata, johon kuuluu monia perinteisiä alkueläimet, kuten Apicomplexa, ripsieläimet ja panssarisiimalevät, muodostaa haaran kanssa chromista, mutta ne kuitenkin sijoitetaan alkueläin valtakunnan. Näin ollen chromista eivät ole holophyletic. Liikuttaminen alveolata uuteen valtakunta nimeltään chromalveolata ratkaisisi tämän ongelman. Tämä johtaisi todennäköisesti ristiriitoja ja sekaannusta, sillä monet alveolata on pitkään pidetty alkueläimet.
Tulane University (©2000).
Comments are closed, but trackbacks and pingbacks are open.